
 French
French Deutsch
DeutschPliocene
This article needs additional citations for verification. (January 2019) |
| Pliocene | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Chronology | |||||||||||
| |||||||||||
| Etymology | |||||||||||
| Name formality | Formal | ||||||||||
| Usage information | |||||||||||
| Celestial body | Earth | ||||||||||
| Regional usage | Global (ICS) | ||||||||||
| Time scale(s) used | ICS Time Scale | ||||||||||
| Definition | |||||||||||
| Chronological unit | Epoch | ||||||||||
| Stratigraphic unit | Series | ||||||||||
| Time span formality | Formal | ||||||||||
| Lower boundary definition | Base of the Thvera magnetic event (C3n.4n), which is only 96 ka (5 precession cycles) younger than the GSSP | ||||||||||
| Lower boundary GSSP | Heraclea Minoa section, Heraclea Minoa, Cattolica Eraclea, Sicily, Italy 37°23′30″N 13°16′50″E / 37.3917°N 13.2806°E | ||||||||||
| Lower GSSP ratified | 2000[4] | ||||||||||
| Upper boundary definition |
| ||||||||||
| Upper boundary GSSP | Monte San Nicola Section, Gela, Sicily, Italy 37°08′49″N 14°12′13″E / 37.1469°N 14.2035°E | ||||||||||
| Upper GSSP ratified | 2009 (as base of Quaternary and Pleistocene)[5] | ||||||||||
| Part of a series on |
| Human history and prehistory |
|---|
| ↑ before Homo (Pliocene epoch) |
| ↓ Future (Holocene epoch) |
The Pliocene ( /ˈplaɪ.əsiːn, ˈplaɪ.oʊ-/ PLY-ə-seen, PLY-oh-;[6][7] also Pleiocene)[8] is the epoch in the geologic time scale that extends from 5.333 million to 2.58[9] million years ago. It is the second and most recent epoch of the Neogene Period in the Cenozoic Era. The Pliocene follows the Miocene Epoch and is followed by the Pleistocene Epoch. Prior to the 2009 revision of the geologic time scale, which placed the four most recent major glaciations entirely within the Pleistocene, the Pliocene also included the Gelasian Stage, which lasted from 2.588 to 1.806 million years ago, and is now included in the Pleistocene.[10]
As with other older geologic periods, the geological strata that define the start and end are well-identified but the exact dates of the start and end of the epoch are slightly uncertain. The boundaries defining the Pliocene are not set at an easily identified worldwide event but rather at regional boundaries between the warmer Miocene and the relatively cooler Pleistocene. The upper boundary was set at the start of the Pleistocene glaciations.
Etymology[edit]
Charles Lyell (later Sir Charles) gave the Pliocene its name in Principles of Geology (volume 3, 1833).[11]
The word pliocene comes from the Greek words πλεῖον (pleion, "more") and καινός (kainos, "new" or "recent")[12] and means roughly "continuation of the recent", referring to the essentially modern marine mollusc fauna.
Subdivisions[edit]

In the official timescale of the ICS, the Pliocene is subdivided into two stages. From youngest to oldest they are:
- Piacenzian (3.600–2.58 Ma)[13]
- Zanclean (5.333–3.600 Ma)[4]
The Piacenzian is sometimes referred to as the Late Pliocene, whereas the Zanclean is referred to as the Early Pliocene.
In the system of
- North American Land Mammal Ages (NALMA) include Hemphillian (9–4.75 Ma),[14][15] and Blancan (4.75–1.6 Ma).[16] The Blancan extends forward into the Pleistocene.
- South American Land Mammal Ages (SALMA) include Montehermosan (6.8–4.0 Ma), Chapadmalalan (4.0–3.0 Ma) and Uquian (3.0–1.2 Ma).[17]
In the Paratethys area (central Europe and parts of western Asia) the Pliocene contains the Dacian (roughly equal to the Zanclean) and Romanian (roughly equal to the Piacenzian and Gelasian together) stages. As usual in stratigraphy, there are many other regional and local subdivisions in use.
In Britain, the Pliocene is divided into the following stages (old to young): Gedgravian, Waltonian, Pre-Ludhamian, Ludhamian, Thurnian, Bramertonian or Antian, Pre-Pastonian or Baventian, Pastonian and Beestonian. In the Netherlands the Pliocene is divided into these stages (old to young): Brunssumian C, Reuverian A, Reuverian B, Reuverian C, Praetiglian, Tiglian A, Tiglian B, Tiglian C1-4b, Tiglian C4c, Tiglian C5, Tiglian C6 and Eburonian. The exact correlations between these local stages and the International Commission on Stratigraphy (ICS) stages is still a matter of detail.[18]
Climate[edit]

The beginning of the Pliocene was marked by an increase in global temperatures relative to the cooler Messinian related to the 1.2 million year obliquity amplitude modulation cycle.[19] The global average temperature in the mid-Pliocene (3.3–3 mya) was 2–3 °C higher than today,[20] carbon dioxide levels were the same as today (400 ppm),[21] and global sea level was 25 m higher.[22] The northern hemisphere ice sheet was ephemeral before the onset of extensive glaciation over Greenland that occurred in the late Pliocene around 3 Ma.[23] The formation of an Arctic ice cap is signaled by an abrupt shift in oxygen isotope ratios and ice-rafted cobbles in the North Atlantic and North Pacific Ocean beds.[24] Mid-latitude glaciation was probably underway before the end of the epoch. The global cooling that occurred during the Pliocene may have accelerated on the disappearance of forests and the spread of grasslands and savannas.[25]
Paleogeography[edit]

Continents continued to drift, moving from positions possibly as far as 250 km from their present locations to positions only 70 km from their current locations. South America became linked to North America through the Isthmus of Panama during the Pliocene, making possible the Great American Interchange and bringing a nearly complete end to South America's distinctive native ungulate fauna,[26] though other South American lineages like its predatory mammals were already extinct by this point and others like xenarthrans continued to do well afterwards. The formation of the Isthmus had major consequences on global temperatures, since warm equatorial ocean currents were cut off and an Atlantic cooling cycle began, with cold Arctic and Antarctic waters dropping temperatures in the now-isolated Atlantic Ocean.[27]
Africa's collision with Europe formed the Mediterranean Sea, cutting off the remnants of the Tethys Ocean. The border between the Miocene and the Pliocene is also the time of the Messinian salinity crisis.[28][29]
During the Late Pliocene, the Himalayas became less active in their uplift, as evidenced by sedimentation changes in the Bengal Fan.[30]
The land bridge between Alaska and Siberia (Beringia) was first flooded near the start of the Pliocene, allowing marine organisms to spread between the Arctic and Pacific Oceans. The bridge would continue to be periodically flooded and restored thereafter.[31]
Pliocene marine formations are exposed in northeast Spain,[32] southern California,[33] New Zealand,[34] and Italy.[35]
During the Pliocene parts of southern Norway and southern Sweden that had been near sea level rose. In Norway this rise elevated the Hardangervidda plateau to 1200 m in the Early Pliocene.[36] In Southern Sweden similar movements elevated the South Swedish highlands leading to a deflection of the ancient Eridanos river from its original path across south-central Sweden into a course south of Sweden.[37]
Environment and evolution of human ancestors[edit]
The Pliocene is bookended by two significant events in the evolution of human ancestors. The first is the appearance of the hominin Australopithecus anamensis in the early Pliocene, around 4.2 million years ago.[38][39][40] The second is the appearance of Homo, the genus that includes modern humans and their closest extinct relatives, near the end of the Pliocene at 2.6 million years ago.[41] Key traits that evolved among hominins during the Pliocene include terrestrial bipedality and, by the end of the Pliocene, encephalized brains (brains with a large neocortex relative to body mass[42][a] and stone tool manufacture.[43]
Improvements in dating methods and in the use of climate proxies have provided scientists with the means to test hypotheses of the evolution of human ancestors.[43][44] Early hypotheses of the evolution of human traits emphasized the selective pressures produced by particular habitats. For example, many scientists have long favored the savannah hypothesis. This proposes that the evolution of terrestrial bipedality and other traits was an adaptive response to Pliocene climate change that transformed forests into more open savannah. This was championed by Grafton Elliot Smith in his 1924 book, The Evolution of Man, as "the unknown world beyond the trees", and was further elaborated by Raymond Dart as the killer ape theory.[45] Other scientists, such as Sherwood L. Washburn, emphasized an intrinsic model of hominin evolution. According to this model, early evolutionary developments triggered later developments. The model placed little emphasis on the surrounding environment.[46] Anthropologists tended to focus on intrinsic models while geologists and vertebrate paleontologists tended to put greater emphasis on habitats.[47]
Alternatives to the savanna hypothesis include the woodland/forest hypothesis, which emphasizes the evolution of hominins in closed habitats, or hypotheses emphasizing the influence of colder habitats at higher latitudes or the influence of seasonal variation. More recent research has emphasized the variability selection hypothesis, which proposes that variability in climate fostered development of hominin traits.[43] Improved climate proxies show that the Pliocene climate of east Africa was highly variable, suggesting that adaptability to varying conditions was more important in driving hominin evolution than the steady pressure of a particular habitat.[42]
Flora[edit]
The change to a cooler, drier, more seasonal climate had considerable impacts on Pliocene vegetation, reducing tropical species worldwide. Deciduous forests proliferated, coniferous forests and tundra covered much of the north, and grasslands spread on all continents (except Antarctica). Tropical forests were limited to a tight band around the equator, and in addition to dry savannahs, deserts appeared in Asia and Africa.[48][failed verification]
Fauna[edit]
Both marine and continental faunas were essentially modern, although continental faunas were a bit more primitive than today.
The land mass collisions meant great migration and mixing of previously isolated species, such as in the Great American Interchange. Herbivores got bigger, as did specialized predators.
Image gallery[edit]
-

-

-
 A gastropod and attached serpulid wormtube from the Pliocene of Cyprus
A gastropod and attached serpulid wormtube from the Pliocene of Cyprus -
 The gastropod Turritella carinata from the Pliocene of Cyprus
The gastropod Turritella carinata from the Pliocene of Cyprus -
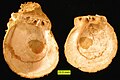
-
 The limpet Diodora italica from the Pliocene of Cyprus
The limpet Diodora italica from the Pliocene of Cyprus -

-
 The gastropod Aporrhais from the Pliocene of Cyprus
The gastropod Aporrhais from the Pliocene of Cyprus -
 The arcid bivalve Anadara from the Pliocene of Cyprus
The arcid bivalve Anadara from the Pliocene of Cyprus -
 The pectenid bivalve Ammusium cristatum from the Pliocene of Cyprus
The pectenid bivalve Ammusium cristatum from the Pliocene of Cyprus -
 Vermetid gastropod Petaloconchus intortus attached to a branch of the coral Cladocora from the Pliocene of Cyprus
Vermetid gastropod Petaloconchus intortus attached to a branch of the coral Cladocora from the Pliocene of Cyprus -













Mammals[edit]
−10 — – −9 — – −8 — – −7 — – −6 — – −5 — – −4 — – −3 — – −2 — – −1 — – 0 — |
| |||||||||||||||||||||||||||||

In North America, rodents, large mastodons and gomphotheres, and opossums continued successfully, while hoofed animals (ungulates) declined, with camel, deer and horse all seeing populations recede. Three-toed horses (Nannippus), oreodonts, protoceratids, and chalicotheres became extinct. Borophagine dogs and Agriotherium became extinct, but other carnivores including the weasel family diversified, and dogs and short-faced bears did well. Ground sloths, huge glyptodonts, and armadillos came north with the formation of the Isthmus of Panama.
In Eurasia rodents did well, while primate distribution declined. Elephants, gomphotheres and stegodonts were successful in Asia (the largest land mammals of the Pliocene were such proboscideans as Deinotherium, Anancus and Mammut borsoni[49]), and hyraxes migrated north from Africa. Horse diversity declined, while tapirs and rhinos did fairly well. Bovines and antelopes were successful; some camel species crossed into Asia from North America. Hyenas and early saber-toothed cats appeared, joining other predators including dogs, bears and weasels.
 |
Africa was dominated by hoofed animals, and primates continued their evolution, with australopithecines (some of the first hominins) and baboon-like monkeys such as the Dinopithecus appearing in the late Pliocene. Rodents were successful, and elephant populations increased. Cows and antelopes continued diversification and overtook pigs in numbers of species. Early giraffes appeared. Horses and modern rhinos came onto the scene. Bears, dogs and weasels (originally from North America) joined cats, hyenas and civets as the African predators, forcing hyenas to adapt as specialized scavengers. Most mustelids in Africa declined as a result of increased competition from the new predators, although Enhydriodon omoensis remained an unusually successful terrestrial predator.
South America was invaded by North American species for the first time since the Cretaceous, with North American rodents and primates mixing with southern forms. Litopterns and the notoungulates, South American natives, were mostly wiped out, except for the macrauchenids and toxodonts, which managed to survive. Small weasel-like carnivorous mustelids, coatis and short-faced bears migrated from the north. Grazing glyptodonts, browsing giant ground sloths and smaller caviomorph rodents, pampatheres, and armadillos did the opposite, migrating to the north and thriving there.
The marsupials remained the dominant Australian mammals, with herbivore forms including wombats and kangaroos, and the huge Diprotodon. Carnivorous marsupials continued hunting in the Pliocene, including dasyurids, the dog-like thylacine and cat-like Thylacoleo. The first rodents arrived in Australia. The modern platypus, a monotreme, appeared.
Birds[edit]

The predatory South American phorusrhacids were rare in this time; among the last was Titanis, a large phorusrhacid that migrated to North America and rivaled mammals as top predator. Other birds probably evolved at this time, some modern (such as the genera Cygnus, Bubo, Struthio and Corvus), some now extinct.
Reptiles and amphibians[edit]
Alligators and crocodiles died out in Europe as the climate cooled. Venomous snake genera continued to increase as more rodents and birds evolved. Rattlesnakes first appeared in the Pliocene. The modern species Alligator mississippiensis, having evolved in the Miocene, continued into the Pliocene, except with a more northern range; specimens have been found in very late Miocene deposits of Tennessee. Giant tortoises still thrived in North America, with genera like Hesperotestudo. Madtsoid snakes were still present in Australia. The amphibian order Allocaudata became extinct.
Corals[edit]
The Pliocene was a high water mark for species diversity among Caribbean corals. From 5 to 2 Ma, coral species origination rates were relatively high in the Caribbean, although a noticeable extinction event and drop in diversity occurred at the end of this interval.[50]
Oceans[edit]
This article needs additional citations for verification. (May 2021) |
Oceans continued to be relatively warm during the Pliocene, though they continued cooling. The Arctic ice cap formed, drying the climate and increasing cool shallow currents in the North Atlantic. Deep cold currents flowed from the Antarctic.
The formation of the Isthmus of Panama about 3.5 million years ago[51] cut off the final remnant of what was once essentially a circum-equatorial current that had existed since the Cretaceous and the early Cenozoic. This may have contributed to further cooling of the oceans worldwide.
The Pliocene seas were alive with sea cows, seals, sea lions and sharks.
See also[edit]
- List of fossil sites (with link directory)
Notes[edit]
- ^ Because of the 2009 reassignment of the Pliocene-Pleistocene boundary from 1.8 to 2.6 million years ago, older papers on Pliocene hominin evolution sometimes include events that would now be regarded as taking place in the early Pleistocene.
References[edit]
- ^ Krijgsman, W.; Garcés, M.; Langereis, C. G.; Daams, R.; Van Dam, J.; Van Der Meulen, A. J.; Agustí, J.; Cabrera, L. (1996). "A new chronology for the middle to late Miocene continental record in Spain". Earth and Planetary Science Letters. 142 (3–4): 367–380. Bibcode:1996E&PSL.142..367K. doi:10.1016/0012-821X(96)00109-4.
- ^ Retallack, G. J. (1997). "Neogene Expansion of the North American Prairie". PALAIOS. 12 (4): 380–390. doi:10.2307/3515337. JSTOR 3515337. Retrieved 11 February 2008.
- ^ "ICS Timescale Chart" (PDF). www.stratigraphy.org.
- ^ a b Van Couvering, John; Castradori, Davide; Cita, Maria; Hilgen, Frederik; Rio, Domenico (September 2000). "The base of the Zanclean Stage and of the Pliocene Series" (PDF). Episodes. 23 (3): 179–187. doi:10.18814/epiiugs/2000/v23i3/005.
- ^ Gibbard, Philip; Head, Martin (September 2010). "The newly-ratified definition of the Quaternary System/Period and redefinition of the Pleistocene Series/Epoch, and comparison of proposals advanced prior to formal ratification" (PDF). Episodes. 33 (3): 152–158. doi:10.18814/epiiugs/2010/v33i3/002. Retrieved 8 December 2020.
- ^ "Pliocene". Merriam-Webster.com Dictionary.
- ^ "Pliocene". Dictionary.com Unabridged (Online). n.d.
- ^ "Pleiocene". Dictionary.com Unabridged (Online). n.d.
- ^ See the 2014 version of the ICS geologic time scale Archived 2014-05-30 at the Wayback Machine
- ^ Ogg, James George; Ogg, Gabi; Gradstein F. M. (2008). The Concise Geologic Time Scale. Cambridge University Press. pp. 150–1. ISBN 9780521898492.
- ^ See:
- Letter from William Whewell to Charles Lyell dated 31 January 1831 in: Todhunter, Isaac, ed. (1876). William Whewell, D. D., Master of Trinity College, Cambridge: An account of his writings with selections from his literary and scientific correspondence. Vol. 2. London, England: Macmillan and Co. p. 111.
- Lyell, Charles (1833). Principles of Geology, …. Vol. 3. London, England: John Murray. p. 53. From p. 53: "We derive the term Pliocene from πλειων, major, and χαινος, recens, as the major part of the fossil testacea of this epoch are referrible to recent species*."
- ^ "Pliocene". Online Etymology Dictionary.
- ^ Castradori, D.; Rio, D.; Hilgen, F. J.; Lourens, L. J. (1998). "The Global Standard Stratotype-section and Point (GSSP) of the Piacenzian Stage (Middle Pliocene)". Episodes. 21 (2): 88–93. doi:10.18814/epiiugs/1998/v21i2/003.
- ^ Tedford, Richard H.; Albright, L. Barry; Barnosky, Anthony D.; Ferrusquia-Villafranca, Ismael; Hunt, Robert M.; Storer, John E.; Swisher, Carl C.; Voorhies, Michael R.; Webb, S. David; Whistler, David P. (31 December 2004). "6. Mammalian Biochronology of the Arikareean Through Hemphillian Interval (Late Oligocene Through Early Pliocene Epochs)". Late Cretaceous and Cenozoic Mammals of North America: 169–231. doi:10.7312/wood13040-008. ISBN 9780231130400.
- ^ Hulbert, Richard C. Jr. (2 August 2016). "Hemphillian North American Land Mammal Age". Fossil Species of Florida. Florida Museum. Retrieved 7 June 2021.
- ^ Hulbert, Richard C. Jr. (2 August 2016). "Blancan North American Land Mammal Age". Fossil Species of Florida. Florida Museum. Retrieved 7 June 2021.
- ^ Flynn, J.; Swisher, C.C. III (1995). "Cenozoic South American Land Mammal Ages: correlation to global geochronology". In William A. Berggren; Dennis V. Kent; Marie-Pierre Aubry; Jan Hardenbol (eds.). Geochronology Time Scales and Global Stratigraphic Correlation. Society for Sedimentary Geology. pp. 317–333. doi:10.2110/pec.95.04.0317.
- ^ Kuhlmann, G.; C.G. Langereis; D. Munsterman; R.-J. van Leeuwen; R. Verreussel; J.E. Meulenkamp; Th.E. Wong (2006). "Integrated chronostratigraphy of the Pliocene-Pleistocene interval and its relation to the regional stratigraphical stages in the southern North Sea region" (PDF). Netherlands Journal of Geosciences. 85 (1): 19–35. Bibcode:2006NJGeo..85...19K. doi:10.1017/S0016774600021405. S2CID 62803118.
- ^ Qin, Jie; Zhang, Rui; Kravchinsky, Vadim A.; Valet, Jean-Pierre; Sagnotti, Leonardo; Li, Jianxing; Xu, Yong; Anwar, Taslima; Yue, Leping (2 April 2022). "1.2 Myr Band of Earth-Mars Obliquity Modulation on the Evolution of Cold Late Miocene to Warm Early Pliocene Climate". Solid Earth. 127 (4). Bibcode:2022JGRB..12724131Q. doi:10.1029/2022JB024131. S2CID 247933545. Retrieved 24 November 2022.
- ^ Robinson, M.; Dowsett, H.J.; Chandler, M.A. (2008). "Pliocene role in assessing future climate impacts". Eos, Transactions, American Geophysical Union. 89 (49): 501–502. Bibcode:2008EOSTr..89..501R. doi:10.1029/2008eo490001.
- ^ "Solutions: Responding to Climate Change". Climate.Nasa.gov. Retrieved 1 September 2016.
- ^ Dwyer, G.S.; Chandler, M.A. (2009). "Mid-Pliocene sea level and continental ice volume based on coupled benthic Mg/Ca palaeotemperatures and oxygen isotopes". Phil. Trans. R. Soc. A. 367 (1886): 157–168. Bibcode:2009RSPTA.367..157D. doi:10.1098/rsta.2008.0222. hdl:10161/6586. PMID 18854304. S2CID 3199617.
- ^ Bartoli, G.; et al. (2005). "Final closure of Panama and the onset of northern hemisphere glaciation". Earth Planet. Sci. Lett. 237 (1–2): 3344. Bibcode:2005E&PSL.237...33B. doi:10.1016/j.epsl.2005.06.020.
- ^ Van Andel (1994), p. 226.
- ^ "The Pliocene epoch". University of California Museum of Paleontology. Retrieved 25 March 2008.
- ^ Webb, S. David (1991). "Ecogeography and the Great American Interchange". Paleobiology. 17 (3): 266–280. Bibcode:1991Pbio...17..266W. doi:10.1017/S0094837300010605. JSTOR 2400869. S2CID 88305955.
- ^ Bartoli, G.; Sarnthein, M.; Weinelt, M.; Erlenkeuser, H.; Garbe-Schönberg, D.; Lea, D.W. (August 2005). "Final closure of Panama and the onset of northern hemisphere glaciation". Earth and Planetary Science Letters. 237 (1–2): 33–44. Bibcode:2005E&PSL.237...33B. doi:10.1016/j.epsl.2005.06.020.
- ^ Gautier, F., Clauzon, G., Suc, J.P., Cravatte, J., Violanti, D., 1994. Age and duration of the Messinian salinity crisis. C.R. Acad. Sci., Paris (IIA) 318, 1103–1109.
- ^ Krijgsman, W (August 1996). "A new chronology for the middle to late Miocene continental record in Spain" (PDF). Earth and Planetary Science Letters. 142 (3–4): 367–380. Bibcode:1996E&PSL.142..367K. doi:10.1016/0012-821X(96)00109-4.
- ^ Chang, Zihan; Zhou, Liping (December 2019). "Evidence for provenance change in deep sea sediments of the Bengal Fan: A 7 million year record from IODP U1444A". Journal of Asian Earth Sciences. 186. Bibcode:2019JAESc.18604008C. doi:10.1016/j.jseaes.2019.104008. S2CID 202902163. Retrieved 6 July 2023.
- ^ Gladenkov, Andrey Yu; Oleinik, Anton E; Marincovich, Louie; Barinov, Konstantin B (July 2002). "A refined age for the earliest opening of Bering Strait". Palaeogeography, Palaeoclimatology, Palaeoecology. 183 (3–4): 321–328. Bibcode:2002PPP...183..321G. doi:10.1016/S0031-0182(02)00249-3. Retrieved 6 July 2023.
- ^ Gibert, Jordi Maria de; Martinell, Jordi (January 1995). "Sedimentary substrate andtrace fossil assemblages in marine Pliocene deposits in Northeast Spain". Geobios. 28: 197–206. Bibcode:1995Geobi..28R.197G. doi:10.1016/S0016-6995(95)80166-9.
- ^ Deméré, Thomas A. (1983). "The Neogene San Diego basin: a review of the marine Pliocene San Diego formation". Cenozoic Marine Sedimentation, Pacific Margin. Pacific Section,m Society for Sedimentary Geology. pp. 187–195. Retrieved 7 June 2021.
- ^ Saul, G.; Naish, T.R.; Abbott, S.T.; Carter, R.M. (1 April 1999). "Sedimentary cyclicity in the marine Pliocene-Pleistocene of the Wanganui basin (New Zealand): Sequence stratigraphic motifs characteristic of the past 2.5 m.y." Geological Society of America Bulletin. 111 (4): 524–537. Bibcode:1999GSAB..111..524S. doi:10.1130/0016-7606(1999)111<0524:SCITMP>2.3.CO;2. Retrieved 6 July 2023.
- ^ Selli, Raimondo (September 1965). "The Pliocene-Pleistocene boundary in Italian marine sections and its relationship to continental stratigraphies". Progress in Oceanography. 4: 67–86. Bibcode:1965PrOce...4...67S. doi:10.1016/0079-6611(65)90041-8.
- ^ Japsen, Peter; Green, Paul F.; Chalmers, James A.; Bonow, Johan M. (17 May 2018). "Mountains of southernmost Norway: uplifted Miocene peneplains and re-exposed Mesozoic surfaces". Journal of the Geological Society. 175 (5): 721–741. Bibcode:2018JGSoc.175..721J. doi:10.1144/jgs2017-157. S2CID 134575021.
- ^ Lidmar-Bergström, Karna; Olvmo, Mats; Bonow, Johan M. (2017). "The South Swedish Dome: a key structure for identification of peneplains and conclusions on Phanerozoic tectonics of an ancient shield". GFF. 139 (4): 244–259. Bibcode:2017GFF...139..244L. doi:10.1080/11035897.2017.1364293. S2CID 134300755.
- ^ Rozé, Simon (28 August 2019). "Le plus vieux crâne fossile d'australopithèque a été découvert en Éthiopie". RFI (in French). Retrieved 18 February 2023.
- ^ Haile-Selassie, Y (27 October 2010). "Phylogeny of early Australopithecus: new fossil evidence from the Woranso-Mille (central Afar, Ethiopia)". Philosophical Transactions of the Royal Society B: Biological Sciences. 365 (1556): 3323–3331. doi:10.1098/rstb.2010.0064. PMC 2981958. PMID 20855306.
- ^ Lewis, Barry; et al. (2013). Understanding Humans: Introduction to Physical Anthropology and Archaeology (11th ed.). Belmont, CA: Wadsworth Publishing.
{{cite book}}: CS1 maint: location missing publisher (link) - ^ Stringer CB (1994). "Evolution of early humans". In Jones S, Martin R, Pilbeam D (eds.). The Cambridge Encyclopedia of Human Evolution. Cambridge: Cambridge University Press. p. 242.
- ^ a b Potts, R. (2007), Bobe, René; Alemseged, Zeresenay; Behrensmeyer, Anna K. (eds.), "Environmental hypotheses of Pliocene human evolution", Hominin Environments in the East African Pliocene: An Assessment of the Faunal Evidence, Vertebrate Paleobiology and Paleoanthropology Series, Dordrecht: Springer Netherlands, pp. 25–49, doi:10.1007/978-1-4020-3098-7_2, ISBN 978-1-4020-3098-7
- ^ a b c Potts, Richard (3 January 1999). "Environmental hypotheses of hominin evolution". American Journal of Biological Anthropology. 107 (S27): 93–136. doi:10.1002/(SICI)1096-8644(1998)107:27+<93::AID-AJPA5>3.0.CO;2-X. PMID 9881524.
- ^ deMenocal, Peter B (3 March 2004). "African climate change and faunal evolution during the Pliocene–Pleistocene". Earth and Planetary Science Letters. 220 (1): 3–24. Bibcode:2004E&PSL.220....3D. doi:10.1016/S0012-821X(04)00003-2. ISSN 0012-821X.
- ^ Dart, R (1953). "The predatory transition from ape to man". Int. Anthrop. Ling. Rev. 1: 201–218.
- ^ Washburn, SL (1960). "Tools and human evolution". Sci. Am. 203 (3): 63–75. Bibcode:1960SciAm.203c..62W. doi:10.1038/scientificamerican0960-62. JSTOR 24940615. PMID 13843002.
- ^ Potts 1999, pp. 106–108.
- ^ Mares, Micheal A., ed. (1999). "Miocene". Encyclopedia of Deserts. University of Oaklahoma Press. ISBN 0-8061-3146-2.
- ^ Charles A. Repenning, Richard H. Tedford (2013). Fossils of the Carpathian Region. Indiana University Press. p. 373. ISBN 9780253009876. Retrieved 23 August 2022.
- ^ Budd, A. F. (April 2000). "Diversity and extinction in the Cenozoic history of Caribbean reefs". Coral Reefs. 19: 25–35. doi:10.1007/s003380050222. S2CID 1062406. Retrieved 10 May 2023.
- ^ Keigwin, Lloyd D. (1 October 1978). "Pliocene closing of the Isthmus of Panama, based on biostratigraphic evidence from nearby Pacific Ocean and Caribbean Sea cores". Geology. 6 (10): 630–634. Bibcode:1978Geo.....6..630K. doi:10.1130/0091-7613(1978)6<630:PCOTIO>2.0.CO;2. ISSN 0091-7613.
{kind=link}
{kind=link}
Further reading[edit]
- Comins, Niel F.; William J. Kaufmann III (2005). Discovering the Universe (7th ed.). New York, NY: Susan Finnemore Brennan. ISBN 978-0-7167-7584-3.
- Gradstein, F.M.; Ogg, J.G. & Smith, A.G.; 2004: A Geologic Time Scale 2004, Cambridge University Press.
- Ogg, Jim (June 2004). "Overview of Global Boundary Stratotype Sections and Points (GSSP's)". Archived from the original on 23 April 2006. Retrieved 30 April 2006.
- Van Andel, Tjeerd H. (1994). New Views on an Old Planet: a History of Global Change (2nd ed.). Cambridge: Cambridge University Press. ISBN 978-0-521-44243-5.
External links[edit]
- Mid-Pliocene Global Warming: NASA/GISS Climate Modeling
- Palaeos Pliocene
- PBS Change: Deep Time: Pliocene
- Possible Pliocene supernova
- "Supernova dealt deaths on Earth? Stellar blasts may have killed ancient marine life" Science News Online retrieved February 2, 2002
- UCMP Berkeley Pliocene Epoch Page
- Pliocene Microfossils: 100+ images of Pliocene Foraminifera
- Human Timeline (Interactive) – Smithsonian, National Museum of Natural History (August 2016).